Yoël Forterre1, Philippe Marmottant2, Catherine Quilliet2 ja Xavier Noblin3 – DOI: 10.1051/epn/2016104
Taimed liiguvad mitte ainult tuule mõjul või kasvades. Kuigi neil ei ole lihaseid, on mõnedel taimedel välja arenenud mehhanismid, mille abil nad saavad teha hämmastavalt kiireid liigutusi (kiirused ulatuvad kuni 10 m/s) ja kiirendusi (tuhandeid g-sid), mis on võrreldavad loomade liigutuste kiirusega ja isegi ületavad neid.
Neid kiireid liikumisi vajavad taimed elutähtsate funktsioonide täitmiseks, nagu seda on paljunemine (mõnede orhideede õietolmu katapulteerumine, seemnete plahvatuslik väljutamine), kaitse kahjurite vastu (häbeliku mimoosil (Mimosa pudica) kokkuvoltimine) ja toitumine (loomtoiduliste taimede püünised). Millised on taimede selliseid kiireid liikumisi tekitavad füüsikalised mehhanismid. Artikli aluseks on hiljutised uuringud, mis on läbi viidud füüsika ja bioloogia piirimail.
Vee põhjustatud liikumised: siserõhk, osmoos ja aurumine
Esimese asjana paneme tähele, et erinevalt loomarakkudest on taimerakud ümbritsetud peamiselt tselluloosist koosnevate jäikade rakukestadega [1]. See takistab taimerakkudes, kokkutõmbumisvõimeliste valkude kasutamist nagu see toimub loomade lihasrakkudes. See-eest võimaldavad taimerakkude jäigad seinad hoida väga kõrget vee siserõhku, mis on tüüpiliselt 4-8 baari ja erilistes rakkudes isegil kuni 40 baari. Veel võib taimedes olla ka negatiivne rõhk (alarõhk), mille põhjustab kohesioonijõud vee molekulide vahel. Sellise rõhu puhul tõmbuvad raku seinad sissepoole, sealjuures takistab puittaimede täielikult välja arenenud rakukest struktuuri kahjustumist isegi kõrgeimate alarõhkude korral (~ 100 baari).
Füüsikaliset on sellised kõrged hüdrostaatilise rõhu absoluutvärtused põhjustatud osmoosist ja aurustumisest [2,3]. Vee liikumine, voog taime ja seda ümbritseva keskkonna vahel kutsub esile taimede rakkude paisumise või kokkutõmbumise. See põhjustab siserõhu, mis omakorda põhjustab silmaga nähtavaid mehaanilisi liigutusi. Mõned sellised liigutused on põhjustatud pöördumatust rakukestade deformatsioonist, nagu näiteks kasvamine ja valguse poole suunatud või gravitatsiooni järgi orienteeritud liikumine (Joonis 1a). Mõnedel teistel juhtudel on tegemist pööratavate liikumistega, nagu näiteks häbeliku mimoosi tundlike lehtede osmootiliselt indutseeritud kokkuvoltimine (Joonis 1c), või männikäbide avanemine/sulgumine vastavalt õhuniiskusele (Joonis 1b).
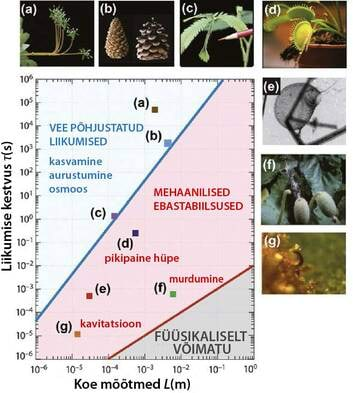
|

|
|---|---|
| Joonis 1 Taimede liigutuste füüsikaline klassifikatsioon liigutuste kestuse τ ja tüüpilise liigutustesse haaratud koe suuruse L järgi (artikli [4] järgi). Piirjoon τ = τp (sinine joon) annab poor- elastse aja (st kiireima liigutuse, mis on võimalik ainult vee transportimisel), ja on arvutatud kasutades diffusiooni koefitsienti D ~ 10-8 m²/s, mis on keskmine väärtus vee liikumisel taimede kudedes. Piirjoon τ = τi (punane joon) annab inertsiaalse aja, st kiireima võimaliku liikumise, kui rakenduvad inerts ja elastsus, ja on arvutatud kasutades tiheduse väärtust ρ = 1000 kg/m3 ja Youngi moodulit E = 10 MPa. (a) Kasvu vastus gravitatsioonile (geotropism). (b) Männi käbisoomuste liikumine vastavalt õhuniiskuse muutustele. (c) Häbeliku mimoosi lehtede osmoosist põhjustatud kokkuvoltimine. (d) Loomtoiduline taim kärbsepüünis. (e) Loomtoiduline taim vesihernes (Utricularia) perekonnast (f) Pritskurgi (Ecballium elaterium) seemnete väljapaiskumine (g) Sõnajala eosla katapuldi-laadne liikumine. | Niimoodi toimub häbeliku mimoosi lehtede kiire kokkuvoltimine. |
Vee põhjustatud liigutuste kiiruse füüsikalised piirid: poor-elastne ajaskaala
Ainult vee põhjustatud liigutuste kiirus on piiratud vee voolamise kiirusega kudedes. [4]. Poorses ja elastses keskkonnas, nagu seda on taime kude, on voolamine difusiivne protsess, mis toimub karakteristliku aja τp=L/D jooksul, kus L on vee liikumise ulatus ja D on efektiivne difusiooni koefitsient. Seda aega τp, nimetatakse „poor-elastseks ajaks“ (poroelastic time), ja joonisel 1 on seda võrreldud taimeriigis toimuvate liigutuste kestustega τ ja liigutusse haaratud koe suurusega L. Tõepoolest, liikumised mis on põhjustatud kasvamisest (Joonis. 1a) või õhuniiskuse muutumisega (Joonis 1b) paiknevad sellel graafikul ülevalpool piirjoont τ = τp ja see on kooskõlas nende puhtalt „hüdraulilise“ iseloomuga. Ka häbeliku mimoosi lehe kiire kokkuvoltimine kuulub sellese rühma - ehkki liigutuse on väga kiire (tüüpiliselt 1 s) on väiksed ka liikumisse haaratud koe mõõtmed.
Taimede liigutuste kiirenemine: elastsus ja mehaaniline ebastabiilsus
Kui taimed kasutaksid liigutustes vaid vee transporti, ei saaks nende liikumised olla kiiremad, kui seda määrab poor-elastne aeg. Samas joonis 1 näitab, et paljud taimed siiski suudavad seda hüdrodünaamilist piiri ületada. Sellised liigutused kasutavad mehaanilisi ebastabiilsusi, st varasemalt salvestatud energia kiiret vabastamist. Taimede juures saab eristada kaht tüüpi ebastabiilsusi: (i) pikipaindest põhjustatud ebastabiilsused, ja (ii) tahke aine või vedeliku purse seemnete või pooride väljapaiskumine. Vaatleme neid kaht mehhanismi lähemalt.
|

|
|---|---|
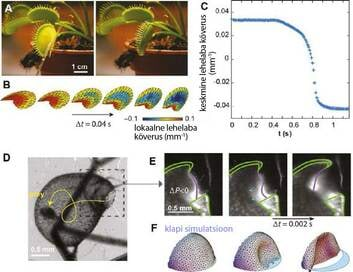
| Joonis 2 Pikipaine ebastabiilsus kärbsepüünises (a-c) ja vesihernes (d-f). (A) kärbsepüünis on avatud (pilt vasakul) ja suletud (pilt paremal). (B) Sulguva lehelaba kuju 3D mudel (eksperimendi andmed, ?t on aeg kahe järjestikuse pildi vahel). (C) Ruumiliselt keskmistatud lehelaba kumeruse sõltuvus ajast. Püünis sulgus ajahetkel t = 0. (D) Vesihernes püünis ja väikeste vähiliste trajektoor, mis imeti püünisesse pärast sulguri avanemist. (E) Sulguri kuju muutumine avanemisel, nähtuna fluorestsents-mikroskoobis. (F) Sulguri avanemise arvutisimulatsioon. (joonised A-C on artikli [5] järgi; joonised D-F on artikli [7] järgi) | Kärbsepüünis oskab oma lehelabadest moodustunud püünist väga kiiresti kokku tõmmata. |
Pikipaine ebastabiilsus: kärbsepüünis and vesihernes
Kärbsepüünis koosneb kahest alusele kinnitunud lehelabast (Joonis. 2A). Iga lehelaba sisemine külg sisaldab tundlikke karvakesi, mis tekitavad elektrilise signaali, kui neid puudutatakse. Püünis sulgub mõne sekundi kümnendiku jooksul, nii et selline liigutus ei saa olla põhjustatud vaid vee liikumisest (Joonis 1). Hiljutised uurimused on näidanud, et sulgumise kiirust suurendav mehhanism on ebastabiilne pikipaine – lehed käituvad sarnaselt, kui plõksuga ümber randme keerduv helkur-käevõru [5,6]. Taime kaks lehelaba on avatud asendis kõverdunud väljapoole ja suletud asendis sissepoole (Joonis 2B,C). Enne sulgumist lehelabad „püüavad“ kõverduda sissepoole. Aga kuna oma kumera geomeetrilise kuju tõttu ei ole see võimalik, siis akumuleerub püünise labadesse elastsusjõu energia. Kui elastsusjõud muutub piisavalt suureks, toimub lehelaba liigutus ühest olekust teise ning püünis sulgub kiiresti.

|
|
|---|---|
| Muu seas näitab video ka seda, kuida pritskurk oma seemneid välja pritsib. | Vesihernes püüab saaki, imedes ennast ümbritseva vee väga kiiresti lehest moodustunud põiekujulisse püünisesse. |
Uurimused on näidanud, et samasugust mehhanismi liigutab vesiherne imemispüünist [7]. Iga selle veetaime püünis on elastne, suletud, põiekujuline leht, kus rõhk aeglaselt väheneb (Joonis 2d). Püünis on hermeetiline tänu painduvale sulgurile, millel paiknevad ka mõned pikad tundlikud karvakesed. Algasendis on põie uks justkui kuppel, mis on kumerdunud väljapoole ning mis sellisena paneb rõhu vähenemisele vastu sarnaselt gooti stiilis kiriku kuplitega. Kui selle kupli tugevusvaru ammendub, kas siis elektrisignaali mõjul või lihtsalt mõne nõrga koha tõttu, pöördub kuppel teisipidi ja avaneb kiiresti (Joonis 2E,F). Kuna püünises on alarõhk imetakse sinna väga kiiresti (vaid ühe millisekundi jooksul) vett. Kiirendus selles protsessis on kuni 600g-d, ja väiksed putukad ei suuda vastu sellist voolu ujuda, sattudes püünisesse. (Joonis 2D).
Kavitatsiooniline ebastabiilsus: eoste väljapaiskumine
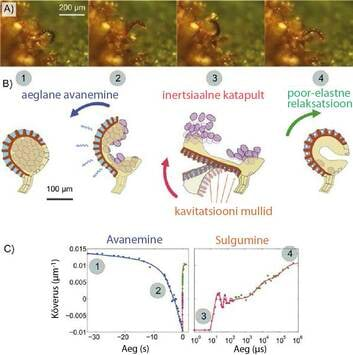
Kiireid liigutusi on vaja ka seemnete, õietolmu ja eoste väljaheitmiseks. Pritskurk (Joonis 1f) on oma seemnete väljaheitmise meetodi pärast hästi tuntud, aga kõige elegantsemat mehhamisi oma eoste väljaheitmiseks kasutab tõenäoliselt sõnajalg (Joonis 3A). Enamustel sõnajala liikidel on eosed sfäärilistes kapslites (eosla), mida ümbritsed kaarekujuline, paljusid veega täidetud rakke sisaldav riba (??) (Joonis 3B). Vee aurumine neist rakkudest tekitab negatiivse rõhu, mis tõmbab raku seinu sissepoole. Kuna raku seinad on ringi välisosas õhemad, saab ring painduda eosla suunas. Nii tekib eosla väljapoole paindumine kuni hetkeni, kui rakkudesse tekivad kavitatsioonimullid (negatiivne rõhk ca 100 baari), mis vabastavad kiiresti kogunenud elastsusjõu energia, eosla sulgub ja heidad selle käigus eosed välja nagu katapuldist.
|

|
|---|---|
| Joonis 3 (A) Sõnajala eosla avanemine ja sulgumine (konkreetne eosla on oma esoed juba välja heitnud). (B) Heite mehhanismi neli etappi, mis vastavad neljale fotole piltidel (A). (C) Eosla keskmise kõveruse sõltuvus ajast, aeglane avanemine (sinine joon) ja kiire sulgumine (punane joon). Logaritmiline skaala aitab märgata kiireid ostsillatsioone sulgumise esimeses etapis ja aeglast poor-elastset sulgumist teises etapis (joonised on artikli [8] järgi). | Sõnajala eoste avanemine ja sulgumine, mille käigus heidetakse eosed välja nagu katapuldist. |
Kirjeldatud mehhanismi on ligikaudselt teatud juba üle saja aasta, aga paljud sellega seotud küsimused on saanud vastused alles hiljuti. Näiteks: kuidas suudab sõnajalg oma eosed nii efektiivselt välja heita ilma, et oleks ühtki liikumist järsult katkestavat takistust või stopperit? Keskaegsetes katapultides kasutati stopperit katapuldi heitja poolel teel peatamiseks, et projektiil lendaks minema enne selle suunamist allapoole. Hiljutised uuringu ülikiirete kaameratega on sellele küsimusele vastuse andnud [8]. Protsessis on kaks faasi, mis toimuvad väga erinevate kiirustega (Joonis 3C). Esimene faas toimub kohe pärast kavitatsiooni ja selle käigus ring osaliselt sulgub mõnekümne mikrosekundi jooksul (see n tõenäoliselt kiireim liigutus, mis taimedel esineb). See inertsiaalne faas on nii kiire, et vesi rõnga poorsetes seintes ei jõus ümber paikneda. Selle tõttu vee rõhk järsult tõuseb ja rõnga sulgumise peatub poolel teel, võimaldades ühtlasi eoste katapulteerimise kiirustega rohkem kui 10 m/s. Teises faasis vee rõhk aeglaselt väheneb ja eosla sulgub täielikult mõnesaja millisekundi jooksul. See on hämmastav, kuidas selline väike, veest ja mõnest rakust koosnev süsteem suudab täita kõiki katapuldi olulisi funktsioone, alustades katapuldi vinnastamisest (vee kohesioon ja aurumine) ja vabastamisest (kavitatsioon) ning lõpetades rakkude seinte poor-elastsetest omadustest põhjustatud äkilise pidurdamisega.
Taimed on hüdraulilised süsteemid ja enamik nende liigutustest on lihtsalt osmoosi ja aurumise põhjustatud vee liikumise tulemus. Selliste liigutuste kiirus on seega piiratud vee difusiooni kiirusega taimede kudedes. Selles artiklis me näitasime, kuidas taimed kasutavad mehaanilisi ebastabiilsusi selleks, et kiiresti vabastada pikema aja jooksul salvestatud energia nii, et tekkivate liikumiste kiirus ületab oluliselt poor-elastse piiri. Selliste liigutuste kiirus on piiratud massi kiirendamiseks kuluva ajaga [4], mis määratakse elastsuslainete levimise kiirusega kehas, τi = L/c, kus L on keha suurus ning c=(E/ρ)1/2 on elastsuslainete levimise kiirus kehas ja E on Youngi moodul ning ρ on keha tihedus. Ei ole leitud ühtki taime, mille liigutuse ületaksid seda piiri (Joonis 1).
Kiired taimed ei ole ainult looduse imed. Koos biomimeetika arenguga on nad lootustandev inspiratsiooniallikas tehislike mikrosüsteemide loomiseks, olgu nendeks siis mikro-vedelik täiturid [10], kiiresti toimivad pinnad [11] või hüppavad robotid [12].
Viited:
[1] L. Taiz and E. Zeiger, Plant Physiology, Sunderland MA, Sinauer Assoc. 3rd ed. (2002).
[2] J. Dumais and Y. Forterre, Annu. Rev. Fluid Mech. 44, 453 (2012).
[3] Y. Forterre, J. Exp. Bot. 64, 4745 (2013).
[4] J. M. Skotheim and L. Mahadevan, Science 308, 1308 (2005).
[5] Y. Forterre et al, Nature 433, 421 (2005).
[6] S. Poppinga and M. Joyeux, Phys. Rev. E 84, 041928 (2011).
[7] O. Vincent et al. Proceedings of the Royal Society B 278, 2909 (2011).
[8] X. Noblin et al, Science 335, 1322 (2012)
[9] Llorens C et al., J. R. Soc. Interface 13: 20150930 (2016)
[10] D. Kim and D. J. Beebe, Lab Chip 7, 193 (2007)
[11] D. P. Holmes and A. J. Crosby, Adv. Mater. 19, 3589 (2007).
[12] H. Lee et al, Soft Matter 6, 4342 (2010)
1 Aix-Marseille Universite´ – CNRS UMR 7343, IUSTI – 13453 Marseille Cedex 13, France
2 Universit´e Grenoble Alpes – CNRS UMR 5588, LiPhy – 38402 Saint-Martin-d’H`eres, France
3 Laboratoire de Physique de la Mati`ere Condens´ee – Universit´e Nice Sophia Antipolis CNRS UMR 7336 – 06108 Nice Cedex 2